遺伝子発現の制御:クロマチン、タンパク質
真核生物の遺伝子発現調節
ヒトの場合、大腸菌と比べて栄養素を作るハウスキーピング遺伝子が少ない。これは食物から栄養を摂取できるためであり、必須アミノ酸や必須脂肪酸は体内で合成されない。(体内で合成されない)
真核多細胞生物の遺伝子には
①分化した細胞の機能を発揮する分化遺伝子
②ホメオタシスに関する遺伝子
③発生に関する遺伝子
などがある。
ホメオタシスとは「ストレスになりうる外界の環境変化に対し、生体を安定した状態に保とうとする仕組み」である。哺乳類では神経、免疫、内分泌(ホルモン)などの相互作用で維持している。
クロマチン構造には真性クロマチンと異性クロマチンがある。
・真性クロマチン
転写が活発
・異性クロマチン
構成的ヘテロクロマチン:常に密に凝縮しており、発現していない
条件的ヘテロクロマチン:必要なときに発現する
クロマチンの構造変化での制御
・メチル化
哺乳類の遺伝子ではCpGの配列のC(シトシン)の約70%がメチル化している。塩基のメチル化は真核生物でも原核生物でも行われている。メチル化すると制限酵素で切断できなくなる。修飾は酵素などによって起こる可逆的な反応である。
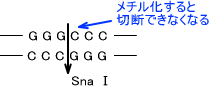
これらの修飾は「エピジェネティックな変化」という。メチル化はDNAの変化であるが、塩基配列の変化ではない。病原体などのほかの生物の外来遺伝子と自分の遺伝子の区別はメチル化の違いによって区別する。
メチル化されている領域はメチル化によって発現が抑えられている。つまり、発現が抑制されている遺伝子は多く、発現していない領域のCpGの80%以上がメチル化している。メチル化はヘテロクロマチン部分に多い。
ハウスキーピング遺伝子はGC配列を多く含むが、この遺伝子領域がメチル化されているために発現しなければ都合が悪い。そのため、この領域はメチル化されていない。この部分はプロモーターやエンハンサーなどがあり、GCアイランド(GCを多く含む配列)を形成している。
細胞によっては同じ遺伝子でもメチル化を受けているものとそうでないものが存在する。例えば、ヘモグロビン遺伝子は赤血球になる細胞以外では発現していない。ヘテロクロマチン化して不活性化しているのである。赤血球にならない細胞ではメチル化の頻度が高い。
・アセチル化
ヒストンはそれ自身にDNAを巻きつけることで、コンパクトに収める役割を果たしている。ヒストンは塩基性タンパク質なのでアミノ基を多く含んでいる。
ヒストンタンパク質のアミノ基がアセチル化すると塩基性が消失する。すると酸性であるDNAとの親和力が低下し、DNAがヒストンから離れやすくなる。
・ゲノムインプリンティング
生殖細胞ができるときはメチル化をなくして初期の状態に戻さないといけない(脱メチル化)。クローン動物の場合では遺伝子を卵細胞に挿入した後で脱メチル化が起こると考えられている。
しかし、クローン動物は老化が早いなどの問題がある。テロメアが短いということがあるが、「初期化が完全でない」「初期化しても正しくメチル化されない」などのメチル化異常も考えられている。
DNA構造の変化による制御
哺乳動物の体細胞は必要ないのに完全な遺伝子セットをもっている。肝臓なら肝臓だけの、心臓なら心臓に必要な遺伝子だけをもつということはないのである。しかし、下等な生物においては遺伝子の数を変化させる場合がある。
・繊毛虫
ゾウリムシの仲間である繊毛虫は小核と大核の二つの核をもっている。小核は全染色体情報をもっており、小核からの必要な遺伝子が増幅されて大核を形成している。mRNAは大核から生合成されている。
・ヒトのリンパ球
ヒトの遺伝子は基本的に変化しない。しかし、リンパ球だけは例外である。多くの抗原に対応するので数万種類以上の抗体を産出する。しかし、一つのリンパ球は一種類の抗体しか作ることができない。
・がん細胞
がん細胞は異常増殖をするので、遺伝子増殖はがん細胞の最も基本的な特徴である。
タンパク質による制御
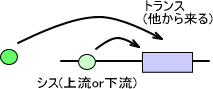
・シスエレメント
シスエレメントは遺伝子の上流または下流に存在し、遺伝子を制御する。シスエレメントには次のようなものがある。
・プロモーター領域:RNAポリメラーゼが結合する
・TATAボックス:転写調節因子が結合する
・CATボックス:転写調節因子が結合する
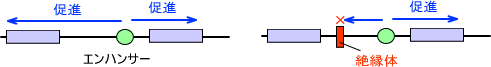
・エンハンサー領域:転写そ促進する塩基配列を持つ
・トランスエレメント
トランスエレメントは、他からやってきてシスエレメントの塩基配列を認識して結合し、遺伝子の転写を調節するタンパク質である。
・エンハンサーの調節
エンハンサーは遠くまで影響を及ぼす。そのままにしておくと無秩序に転写が起こってしまう。そのため、これを防ぐ仕組みが存在する。
・Insulator (絶縁体)
Insulatorが存在することで、それより先のエンハンサーの効果を遮断することができる。
・LCR
会合しているタンパク質と協力して特定の遺伝子発現を制御する。
DNAへのタンパク質の結合
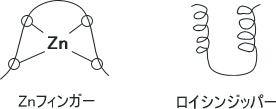
DNAに結合するためにタンパク質は特別な構造をとらないといけない。それらはへリックス-ターン-ヘリックスやZnフィンガー、ロイシンジッパーである。
・ホメオドメイン
動物・植物・菌類の発生を支配している遺伝子領域をホメオボックスと呼ぶ。このホメオボックスによってコードされているDNAに結合するタンパク質部分をホメオドメインという。ホメオドメインタンパク質はヘリックス-ターン-ヘリックスの構造をもつ。
転写の調節
・選択的スプライジング
mRNAhはスプライジングをしてエキソンの部分をつなぎ合わせる。mRNAには複数の遺伝情報があり、エキソンのつなぎあわせ方によって複数のタンパク質ができる場合がある。つまり、必ずしも「1遺伝子=1タンパク質」ではない。
また、作られたタンパク質が切断されて複数のタンパク質ができる場合もある。選択的スプライジングは「転写(mRNAの働き)」に詳しく記述してある。
・mRNAの安定化
mRNAの安定化にはステムループ構造が寄与している。
・miRNA
miRNAは20個程度の塩基配列を持つRNAであり、数百塩基をもつ前駆体のRNAが切断されることによって合成される。miRNAはタンパク質をコードしていない。
miRNAはタンパク質と複合体を形成し、標的のmRNAに結合して翻訳を抑制すると考えられている。(miRNAはmRNAと相補的な配列に結合する)
スポンサードリンク
カテゴリー
スポンサードリンク