転写(mRNAの働き)
RNAはDNAといくつか異なる点があり「一本鎖である」「塩基がチミンでなくウラシルである」「糖がリボースである」などが挙げられる。
主なRNAにはリボソームRNA(rRNA)、トランスファーRNA(tRNA)、メッセンジャーRNA(mRNA)の3種類がある。ここではmRNAについて説明する。
RNAポリメラーゼ
(☆細菌の場合)
RNAの合成はDNAの遺伝情報を元に合成される。また、DNAからRNAへの遺伝情報の転写はRNAポリメラーゼによって行われる。
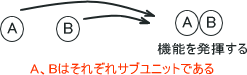
RNAポリメラーゼは2個のαサブユニット、βサブユニット、β´サブユニット、σサブユニットの5個のサブユニットで構成されている。またσ因子は転写開始に関与しており、このRNAポリメラーゼにσ因子が結合することで転写が開始される。
これらのタンパク質は一つのタンパク質ではなく、二つ以上のタンパク質が合わさることで機能を発揮する。
σ因子は解離しやすいため、RNAポリメラーゼが働き出すと離れてしまう。そのためσ因子がはずれたRNAポリメラーゼの形でRNAを合成する。RNAポリメラーゼからσがはずれた酵素をコア酵素といい、RNAポリメラーゼにσ因子がくっついている状態の酵素をホロ酵素という。
σにはいくらかの種類があり、さまざまな菌の状態に対応できるようになっている。

なお、真核生物のRNAポリメラーゼには主な3種類が存在する。
右にその3種類のRNAポリメラーゼを示す。
転写の開始
RNAポリメラーゼによってDNAから合成されるRNAの方向は5´→3´である。転写はDNAのプロモーターという部位にRNAポリメラーゼが結合することで開始する。(プロモーター領域の 1からRNAが作られ始める)
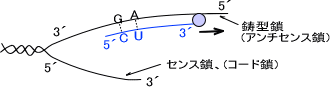
プロモーター領域には共通した塩基配列がある。この配列は転写が開始される最初のDNAの塩基から10塩基と35塩基手前にある。転写が開始される塩基対を 1とすると、これらの配列は-10と-35の領域に存在することになる。
この二つの配列にはコンセンサス配列(共通配列)がある。コンセンサス配列とは「他の生物のDNAを比較したときにだいたい似た配列をもち、同じ機能をもつ領域」のことである。-10領域の共通配列は「TATAAT」であり、-35領域の共通配列は「TGTTGACA」である。
なお-10領域と-35領域はセンス鎖に存在し、転写は鋳型鎖を参考にして転写される。なお、-10領域は発見者の名前からPribnow Box(プリブノー・ボックス)ともいう。

真核生物の場合では、転写の開始はより複雑に制御されている。つまり、哺乳類のプロモーター部分は原核生物よりも複雑であり、さまざまなタンパク質が転写開始の上流に結合することで転写開始を制御している。真核生物でこの転写開始を制御しているタンパク質をトランスエレメント(トランス因子)という。
真核生物では原核生物の-10領域によく似ているTATA(タタ)ボックスという領域が存在する。このTATAボックスにTATAボックス結合因子が結合し、さらに他のタンパク質が結合することで転写が開始する。
主にTATAボックスが転写開始位置を決定し、TATAボックスはHogness box(ホグネス・ボックス)とも呼ばれている。
TATAボックスに結合するタンパク質をもう少し詳しく説明する。まずTBP(TATABox Biding Protein)がDNAに結合する。続いて、種々のタンパク質(TAFというタンパク質)が結合する。これらの結合物をTFⅡDという。
TATAボックスの他にCAATボックスやGCボックスという領域が存在する。これらの配列は全てプロモーター付近に存在しているが、転写開始位置を決定するTATAボックスに対してCAATボックスやGCボックスは転写の頻度を決定する。
真核生物のプロモーター領域に存在する配列
・TATAボックス(Hogness box)
→転写開始位置を決定する。
・CAATボックス、GCボックス
→転写頻度を決定する。
さらに、遺伝子には転写の活性を調節する塩基配列が存在し、これらの配列はエンハンサーまたはサイレンサーと呼ばれている。エンハンサー領域は転写を活性化し、サイレンサー領域は転写を抑制させる。
これらの塩基配列の位置は一定ではなく、転写開始位置の上流(-方向)にも下流( 方向)にも存在する。これらの領域にタンパク質が結合することで効果を発揮する。また、これらの領域にホルモンレセプター蛋白を介して結合するホルモンも存在する。(ex、ステロイドホルモン)
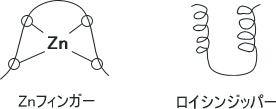
また、DNAに結合するためにタンパク質は特別な構造をとらないといけない。それらはへリックス-ターン-ヘリックスやZnフィンガー、ロイシンジッパーである。
一本のmRNAから作られるタンパク質が一つだけならそのmRNAはモノシストロニックmRNAであり、一本のmRNAから複数のタンパク質が合成されるならそのmRNAはポリシストロニックmRNAである。真核生物のmRNAはモノシストロニックmRNAであることが多く、原核生物のmRNAはポリシストロニックmRNAであることが多い。
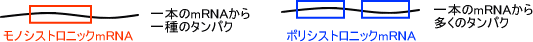
転写の終了
転写開始点としてプロモーターが存在するということを述べた。そして、開始点があるということは転写の終了点もまた存在するということである。この転写終了を示す遺伝子領域をターミネーターという。
RNA合成の終了はDNAの特殊な塩基配列の部分で終了する。この配列部分にはGとCの割合が高く、GとCの配列のすぐ後に三つ以上のAが続いている。
なお、下の図で→の向きで「赤色」「青色」の構造をそれぞれ読むと、全く同じ塩基配列であることが分かる。つまり、これらの塩基配列は上から読んでも下から読んでも同じ配列をしているのである。このような塩基配列をパリンドローム(回文)配列、逆方向反復配列)という。
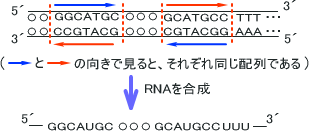
このとき合成されたRNAは一本鎖であるが、GとCの間で強い水素結合が生じるため相手側と塩基対を形成しやすい。そのため、RNAは実際には下のような構造になっている。
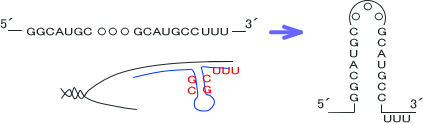
RNA合成は「GCの割合が高い領域がある」という事と「その後すぐ、RNAに3つ以上のUが存在する」という二つの条件がそろうことで転写が終結する。
また転写終結の補助因子としてρ(ロー)因子が存在し、転写終結にρ因子が必要な場合とそうでない場合がある。ρ因子が必要な終結はρ依存性終結であり、必要でない場合はρ非依存性終結である。
RNAの修飾とスプライシング
真核生物のmRNAの5´末端と3´末端は修飾を受ける。DNAは核にあるので、当然ながらmRNAは核で合成される。mRNAはタンパク質生合成をするためにリボソームに移動するとき、安全で抵抗性のある形にしなければならない。そのための修飾である。
まずDNAから合成されて解離したmRNAの5´末端に7-メチルグアノシンが付加する。この5´末端の構造をキャップ構造という。その後、3´末端にポリAポリメラーゼという酵素によってA(アデニン)が200~250個程度くっつけられてmRNAの3´末端を伸長させられる。この3´末端の部分をポリA尾部(Poly A Tail)という。
修飾を受けた後、mRNAはスプライシングを受ける。
多くの真核生物の遺伝子には遺伝情報として翻訳されないDNA塩基配列が存在し、この領域をイントロン(介在配列)という。残りのアミノ酸をコードし、翻訳される領域をエキソンという。
修飾を受けた後のmRNAはエキソンとイントロンの二つの領域が連結して存在している状態である。mRNAはイントロンをなくすためにイントロンの部分を切断し、エキソンの部分をつなぎ合わせる。これがスプライシングである。
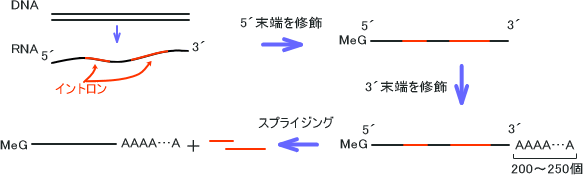
スプライシングは核内で起きており、核内にさまざまな大きさのRNA分子が存在するのはこのためである。このようなさまざまな大きさのmRNAをヘテロ核RNA(hnRNA)という。
スプライシング方法によっては、1本のmRNAからいくつもの異なるエキソンの組み合わせをもつmRNAが生成される。これを選択性スプライシングといい、似ているが異なったタンパク質ができる。
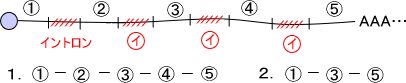
例えば、上の図でイントロン部分だけを切り取ると①-②-③-④-⑤というmRNAができるが、②と④の部分をイントロンと見なして一緒に切り取ると①-③-⑤というmRNAができる。
また、エキソンとイントロンのつなぎ合わせ部分には共通の配列がある。

スプライシングはスプライセオソームが行う。スプライセオソームはRNAとタンパク質から構成されている。つまり、RNA-タンパク質複合体である。
通常、酵素というばタンパク質である。しかし、スプライセオソームの酵素活性の主役はRNAであった。このように酵素作用(触媒活性)をもっているRNAをリボザイムという。
スポンサードリンク
カテゴリー
スポンサードリンク