補体の働きと作用
補体とは
補体は肝臓で合成され、血中に放出される。補体は活性化を受けることによって、その作用を発揮する。
補体は多くの種類がある。英語で補体はcomplementと表し、補体を一般に頭文字をとってCで表す。(Cで表さない補体の種類も存在する)
補体の種類には次のようなものがある。
古典経路 |
第二経路 |
第二経路安定化因子 |
制御系因子 |
|
C1q C1r C1s C4 C2 C3 C5 C6 C7 C8 C9 |
B D |
P C3Nef |
C1INH β1H C4BP |
補体の作用
補体の作用には食細胞による病原菌の食作用の促進や病原菌の細胞膜に孔を空けるなど、病原菌排除に役立っている。
補体の作用には次のようなものがある。
作用 |
関与補体物質 |
|
オプソニン化 |
オプソニン化とは、貪食作用を促進させる作用である。病原菌に補体が結合し、補体レセプターをもつ食細胞が病原菌に結合した補体を認識することで食作用を促進する。 | C4b,C3b |
アナフィラキシー |
肥満細胞(マスト細胞)・好塩基球はヒスタミンを放出させる。ヒスタミンは血管を拡張させて血圧低下を起こす。また、血管透過性亢進も起こす。これによってアレルギーを引き起こす。 | C3a,C5a |
ウイルスの中和 |
外膜をもつウイルスに対して、傷害することにより感染力を失わせる。 | C4b,C3b |
細胞膜破壊 |
補体は最終的に病原菌の細胞膜に孔を作る。これによって、細胞外の物質が細胞内に流入して破裂する。(融解) | C5b678,C5b6789 |
好中球の遊走 |
病原菌に感染した部位に好中球を呼び寄せる作用である。(遊走させる) | C5a,C5b67 |
|
K細胞による ADCC作用の増強 |
抗体と結合した細胞とK細胞との親和性を増強する。(K細胞はADCC作用がある。ADCCとは細胞を傷害する作用のことである。) | C1q,C3b |
|
抗原抗体複合体 可溶化 |
抗原と抗体との再結合を阻止する。これによって抗原抗体複合物は小分子となり、可溶性物質となる。 | C3,C3b,B,D,P |
・抗原抗体複合体可溶化
抗原抗体反応は一方方向の反応ではなく、ある程度の可逆性がある。補体には抗原抗体複合物に結合する能力があり、補体が結合した複合物が抗原と抗体にいったん離れてしまうと、抗原抗体は再び結合しなくなる。
この反応が続いていくと抗原抗体複合物の分子はだんだん小さくなっていく。これによって高分子のために不溶性だった抗原抗体複合物は、可溶性の分子となる。(抗原抗体複合体の可溶化)
不溶性の抗原抗体複合体が蓄積すると、腎炎などさまざまな症状を起こす。補体が作られないヒトでは高分子の抗原抗体となり、自己免疫疾患に陥る。
・貪食の促進
好中球にはFcレセプター,補体レセプターがあり、それぞれ細菌に結合している抗体のFc部や補体(C4b,C3b)を認識すると貪食が促進される。
このときの作用はFcレセプターによる方が強い。補体レセプターは補助作用と考える方が良い。
補体活性化経路
補体が活性化するための代表的な経路は三つあり、それぞれ「古典経路」「第二経路」「レクチン経路」という。
・古典経路
古典経路では、IgMまたはIgGの抗体分子のFc部に補体が結合することから始まる。IgMは1分子でも反応が起こるが、IgGの場合は2分子以上で架橋する必要がある。
抗体に結合する補体はC1qであり、C1qが結合するとC1r酵素活性が活性化してC1sを分解する。これによって、C1sが活性化する。
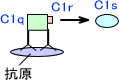
活性化したC1sはC4に作用して、C4aとC4bに分解する。またC1sはC2にも作用して、C2をC2aとC2bに分解する。
C4bは細胞膜に結合できる性質があり、C2aは細胞膜に結合したC4bと結合して複合体を作る。C4bとC2aの複合体はC3を分解するC3転換酵素であり、C3をC3aとC3bに分解する。C3bはC4b2b複合体と結合し、C4b2b3b複合体となる。なお、C3aはアナフェラキシーを引き起こす作用がある。
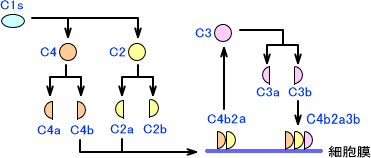
C4b2b3b複合体はC5転換酵素であり、C5をC5aとC5bに分解する。C5bにC6,C7,C8,C9が結合したC5b6789複合体は膜侵襲複合体(membrane attack complex:MAC)と呼ばれ、細胞膜に孔を作って細菌を融解させる。(C5aにはアナフェラキシーを引き起こす作用がある)

古典経路全体の図を下に示す。
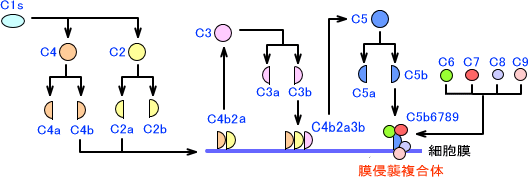
古典経路は最初に抗体が関与する経路である。しかし、他の二つの経路(第二経路、レクチン経路)で抗体の関与はない。
・第二経路
第二経路は代替経路、副経路ともいう。第二経路は微生物の細胞膜上で起こる。
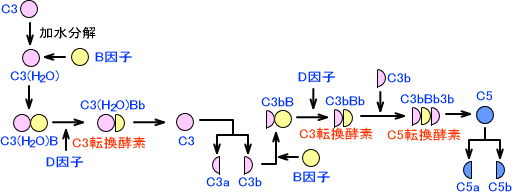
この経路では、まずC3が加水分解を起こしてC3(H2O)となる。C3(H2O)はB因子と結合し、この複合体にD因子が作用するとC3(H2O)Bbとなる。C3(H2O)BbはC3転換酵素である。
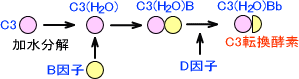
C3(H2O)Bbによって、C3はC3aとC3bに分解される。C3bにB因子が結合してC3bBとなり、これにD因子が作用してC3bBbとなる。C3bBbにもC3転換酵素活性があり、この作用によってC3がどんどん分解される。
C3bBbにC3bが結合すると、今度はC5転換酵素としての作用を示す。C5に作用した後は、古典経路と同じ経路を辿る。
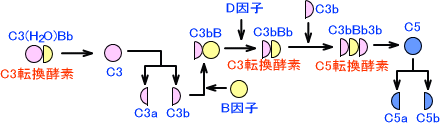
第二経路全体の図を下に示す。
・レクチン経路
微生物の細胞表面にはマンナンやレクチンが存在する。マンナンとは糖であり、D-マンノースからなる多糖である。レクチンは糖と結合する性質をもつタンパクである。
レクチン経路ではマンナン結合タンパク(MLB)とMASP(プロテアーゼの一種)が関与する。MLBとはマンナンと結合する性質のあるタンパクである。
マンナンとMLBが結合するとMASPが活性化する。MASPはC1sのように働き、C4に作用する。後は古典経路と同である。
膜侵襲複合体(MAC)による細胞膜破壊
膜侵襲複合体(MAC)による細胞膜破壊は補体活性化の最終段階である。MACは細胞膜の中に組み込まれ孔が空く。この孔から水やイオンが流入し、細菌が低張となり破裂する。
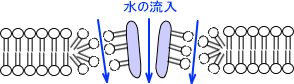
また細胞に孔が開くことによってその周りの構造が変わり、孔の外側からも水・イオンが流入する。
補体の消費
生体内で抗原抗体反応がどんどん起こると、補体もどんどん消費される。主に自己免疫疾患によって補体が消費し、低下する。
補体が低下すると、細菌を排除する能力が低下する。
スポンサードリンク
カテゴリー
スポンサードリンク