自己と非自己の認識
抗原決定基、抗原受容体
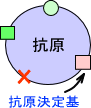
リンパ球が自己と非自己を認識できるのは抗原決定基が存在するからである。リンパ球はこの抗原決定基を認識するのである。
抗原決定基は抗原性の最小単位であり、エピトープともいう。抗原決定基=エピトープ
リンパ球の中でB細胞は細胞表面にある抗原受容体で抗原決定基を認識する。B細胞の抗原受容体なのでBCR(B cell receptor)という。
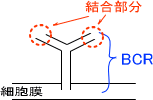
B細胞では抗体分子とほとんど同じ構造のものが細胞表面に出ている。これがBCR(抗原受容体)であり、特定の抗原決定基と結合する性質がある。
BCRと抗原決定基が結合するとB細胞は活性化する。ただし、2つ以上のBCRと抗原が結合して架橋という形をとらないと活性化しない。BCRと抗原が一ヶ所で結合しても活性化は起こらない。

また、一つのB細胞は一種類の抗原受容体しかもっていない。そのため、B細胞が作る抗体も一種類である。
あらゆる抗原に対応するため、B細胞を作り出すときには莫大な数の抗原受容体の中から、その抗原に対応するB細胞クローンだけが選択され増殖する。この考えをクローン説という。
抗原受容体の種類は108個に対応していると言われている。つまり、莫大な数の抗原に対応する抗体産出環境が既に整っているのである。
MHC(主要組織適合遺伝子複合体)
細胞がウイルスに感染するとウイルスは細胞を乗っ取ろうとする。細胞はウイルスに乗っ取られる前に抗原(ウイルス)を切断して、抗原の一部を細胞表面に提示する。T細胞はこの提示された断片を認識するのである。
しかし、抗原だけを提示したのではダメである。抗原はMHC(主要組織適合遺伝子複合体)と結合させて提示しないと意味がない。(MHC 抗原ペプチド)
MHCには2種類あり、それぞれクラスⅠとクラスⅡが存在する。クラスⅠのMHCはキラーT細胞に関与し、クラスⅡのMHCはヘルパーT細胞に関与している。つまり、「MHCクラスⅠ 抗原ペプチド」と「MHCクラスⅡ 抗原ペプチド」は全く別のものである。
クラスⅠとクラスⅡの何が違うかというと、「T細胞上に存在するどのCD抗原と親和性をもつか」が異なっているのである。「血球の細胞表面に存在する抗原分子」をCD(cluster of differentiation)といい、CDにはCD1,CD2 … という具合に何種類も存在する。
キラーT細胞にはCD8があり、ヘルパーT細胞にはCD4がある。
クラスⅠのMHCはCD8と親和性があり、クラスⅡのMHCはCD4と親和性がある。「CD8 T細胞=キラーT細胞」であり、「CD4 T細胞=ヘルパーT細胞」である。

また、クラスⅠのMHCは8~10個の抗原ペプチドを認識し、クラスⅡのMHCは13~17個のペプチドを認識する。
※MHCはHLAとも表現します。
クラスⅠとクラスⅡの抗原提示の仕組み
MHCクラスⅠとMHCクラスⅡでは抗原の取り込み方や処理方法が異なっている。下にクラスⅠとクラスⅡの抗原の処理方法をそれぞれ解説したいと思う。
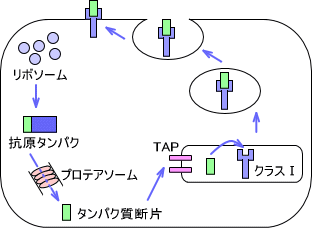
・クラスⅠでの抗原の処理
細胞がウイルスに感染すると、その細胞で合成されるタンパク質はウイルス由来のものへと変わってしまう。MHCクラスⅠで抗原を提示するときは、この合成されたタンパク質抗原を使用する。合成された抗原タンパクはプロテアソームによって小さいペプチドにまで分解される。
ペプチド断片はタンパク質輸送体(TAP)によって小胞体内へ運ばれると、小胞体で合成されたMHCクラスⅠ分子と結合して複合体を形成する。その後、ゴルジ体を経て細胞表面へと運ばれる。
つまり、クラスⅠで抗原を提示するときは細胞内で合成されたペプチドを提示する。
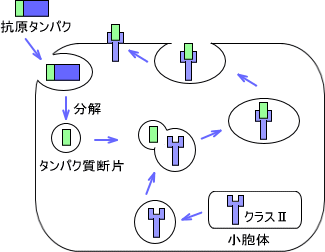
・クラスⅡでの抗原の処理
MHCクラスⅡで提示するための抗原はエンドサイトーシスによって取り込まれる。取り込まれた外来性タンパク質はリソソームによってペプチド断片に分解される。
MHCクラスⅡ分子は小胞体で合成され、他のペプチドとの結合を防ぐためにインバリアント鎖と結合する。クラスⅡ分子がエンドソーム内に入ってインバリアント鎖が切れると、抗原ペプチドと複合体を作る。その後、細胞表面へと運ばれる。
つまり、クラスⅡで抗原を提示するときは細胞外から来たペプチドを提示する。
・T細胞受容体
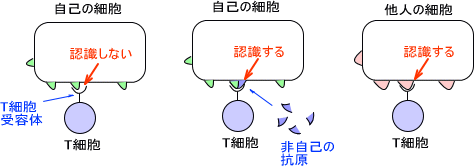
T細胞受容体はT細胞表面に存在する。この受容体は抗原分子(CD)とMHCの複合体に結合する。成熟したキラーT細胞がウイルスに感染した細胞を認識すると感染した細胞は破壊される。
MHCの多様性
T細胞にはさまざまなMHCに噛み合うT細胞受容体(TCR)がある。ただし、自己のMHCに合う受容体だけはもっていない。T細胞は自己だけでは応答しないが、非自己の抗原と合わさると認識する。
普通、遺伝子は父親か母親のどちらかが受け継がれる。しかし、MHCでは父母両方由来の遺伝子が細胞表面に提示される。つまり、MHCには非常に多くの種類が存在する。MHCがこれだけ多型なのには理由がある。
例えば、ヒトのMHCが一種類であるとする。自己のMHCにはT細胞は応答しないので、自己のMHCと全く同じMHCをもつウイルスが現れたとしたら、ヒトの免疫はそのウイルスに対して全く応答しないので、ヒトは絶滅の一途をたどることになる。
こうならないためにMHCは非常に多型となっている。これは動物が生き延びていくのにとても大事なことである。
・拒絶反応の問題
MHCの多様性は、臓器移植の際の拒絶反応の点ではマイナスとなる。前述の通りMHCは多型であり、MHCの構造が似ているヒトは1万人に1人の割合である。細胞表面のMHCが異なるとT細胞はその細胞を攻撃してしまう。これが拒絶反応の起こる仕組みである。
抗原提示細胞(APC)
抗原提示細胞(APC)にはマクロファージ、樹状細胞、B細胞などがある。抗原提示細胞とは、抗原をT細胞に提示することでT細胞を活性化させる細胞のことである。
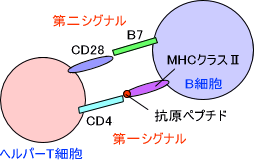
抗原提示細胞はヘルパーT細胞を活性化するとき、抗原提示細胞のMHCクラスⅡとT細胞のCD4を結合させる。これが第一シグナルとなる。しかし、T細胞が活性化するにはもう一回刺激がないといけない。最後に抗原提示細胞のB7とT細胞のCD28が結合し、これが第二シグナルとなってヘルパーT細胞が活性化する。
・マクロファージ
マクロファージは血中では単球とよばれ、抗原をT細胞に提示して活性化させる働きをする。そのため、マクロファージの細胞表面にはFc受容体、補体受容体、MHCクラスⅠ、MHCクラスⅡ分子が存在する。
マクロファージは抗原提示の作用だけではなくウイルス細胞などの粒子を取り込む貪食作用(ファゴサイトーシス)や細胞外液などの溶液を取り込む飲作用(ピノサイトーシス)の作用がある。
・樹状細胞
樹状細胞は皮膚、肺、胃、鼻腔などの生体各組織に広く分布している。樹状細胞はリンパ器官に移行する能力があり、抗原を捕らえると提示できるように分解してリンパ管内へ移動する。移行した後はT細胞に抗原を提示する。
樹状細胞の一番の特徴は「抗原提示細胞の中で最も抗原を提示する能力が高いこと」である。この性質を利用して樹状細胞を取り出して腫瘍の抗原ペプチドを認識させた後、この樹状細胞を再び患者に戻してT細胞を活性化させようという試みがある。これを免疫療法という。
クロスプレゼンテーション
MHCクラスⅠは細胞内で合成された抗原を提示すると述べた。しかし、これには例外がある。それがクロスプレゼンテーションである。
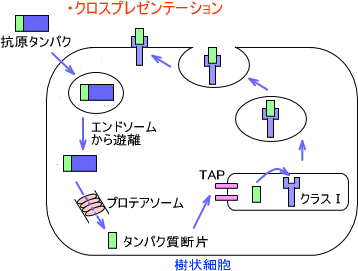
クロスプレゼンテーションは樹状細胞で起こる。この場合、細胞外からの抗原がプロテアソームによって分解され、MHCクラスⅠ分子と結合して細胞表面に提示される。外来の抗原ペプチドをMHCクラスⅠで提示するのである。
スポンサードリンク
カテゴリー
スポンサードリンク