翻訳(tRNAとrRNAの働き)
tRNA
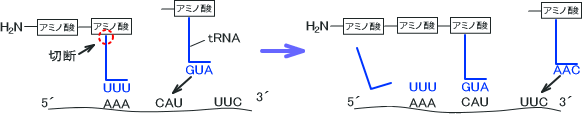
タンパク質の合成はmRNAの配列をもとにリボソームで行われる。tRNAはmRNA中の3つの塩基配列の並び方(コドン)を翻訳することで合成するアミノ酸を決定する。つまり、tRNAはリボソームのタンパク質合成を手伝っている。一つのコドンが一つのアミノ酸に対応している。
なお、tRNAはコドンに対応するアミノ酸を運ぶ役割をしている。
tRNAにはmRNAのコドン(塩基配列の並び方)を認識する領域があり、この領域をアンチコドンという。コドンの情報を読み取ったtRNAのアンチコドンの配列でどのアミノ酸になるかが決定する。コドンの塩基がAであればアンチコドンではUに翻訳され、GならC、CならG、TならAに訳される。
mRNAの塩基配列情報の翻訳はtRNAとアミノアシルtRNAシンテターゼによって行われる。この酵素はATPが必要である。tRNAは一本鎖の核酸であり、tRNAの3´末端にはCCA配列がある。このCCA配列の先にアミノ酸が結合するのだが、アミノ酸をCCAの先に結合させる酵素がアミノアシルtRNAシンテターゼである。。
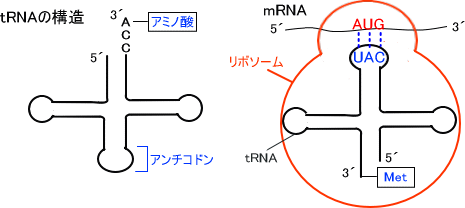
なおtRNAはDNAを転写することで作られるが、tRNAの3´末端のCCA部分に対応する配列は、DNAに存在しないことがある。そして、このtRNAの3´末端のCCA配列はtRNAヌクレオチジルトランスフェラーゼによって付加される。
下にmRNAのそれぞれのコドンに対応するアミノ酸を示す。
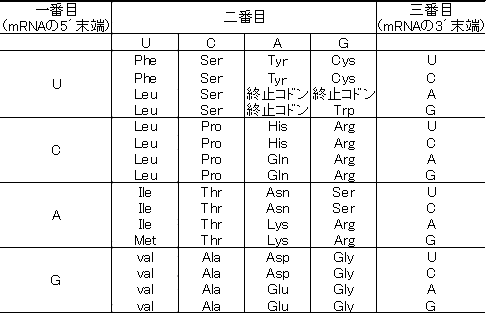
表を見て分かるとおり、mRNAの3番目の塩基はどれでもいい場合がある。この場合はmRNAの3番目に対応するアンチコドンにイノシン(I)を入れるなどして決定権をなくしておく。なお、イノシン(I)はmRNA側のC,U,Aを読むのに有効である。
例えばmRNAの情報がCCA、CCG、CCC、CCTのどの場合でも対応するアミノ酸はPro(プロリン)である。このことを縮重という。
また、このときのアンチコドンの塩基配列がIGGとなっていたとしても、この配列でProに対応できるのである。
DNAでは、それぞれに対応する塩基は決まっていた。しかし、tRNAでは三番目の位置に「この塩基がないといけない」という決まりにある程度のゆとりがある。このゆとりをゆらぎと言い、この仮説をゆらぎ説という。
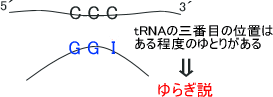
Pro(プロリン)合成コドンである「CCA、CCG、CCC、CCT」の全てに対応するtRNAを合成するには効率が悪い。アンチコドンの5´の位置はI(イノシン)などを用いることで、tRNAの数を減らすようにしている。
なお、ゆとり説の条件下では「G:U」の塩基対が生じることもある。
rRNA(リボソームでのタンパク質合成)
リボソームはタンパク質合成の工場であり、リボソームは数本のRNAとタンパク質から構成されている。このようなリボソームを構成するRNAをrRNAという。つまり、リボソームはrRNAとタンパク質の複合体である。
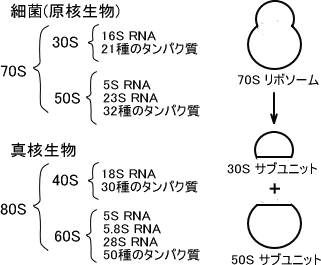
リボソームは二つのサブユニットからできており、その大きさは原核生物と真核生物とでは異なる。原核生物は30Sと50Sのサブユニットから構成され、この二つのサブユニットがくっついた70Sの大きさのリボソームが存在する。真核生物のサブユニットは40Sと60Sから成る80Sのリボソームである。
細菌などの原核生物の30Sサブユニットは16SのrRNA分子と21種類のタンパク質を含み、50Sサブユニットは23Sと5Sの2種類のrRNAと32種類のタンパク質を含んでいる。
真核生物の40Sサブユニットは18SのrRNA分子と30種類のタンパク質を含み、60Sサブユニットは5Sと5.8Sと28Sの3種類のrRNAと50種類のタンパク質を含んでいる。
rRNAを作るDNAは複数箇所に存在する。これによってrRNAの安定供給ができるようになっている。
翻訳の開始
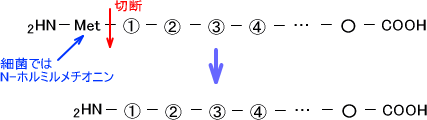
翻訳の開始はMet(メチオニン)から始まる。つまり、翻訳開始のコドンはAUGである。この最初のメチオニンは特殊なメチオニンであり、細菌の場合はN-ホルミルメチオニンである。
細菌での翻訳開始にはまず70Sリボソームが30Sと50Sに解離することから始まる。細菌ではAUGの約10base上流に「AGGAGGU」のShine Dalgarno配列(シャイン・ダルガーノ配列)というコンセンサス配列があり、そこに「30S rRNA、N-ホルモルメチオニンを含むtRNA、Initiation factor(開始因子)、GTP」が結合することで翻訳が開始する。
その後、ATPを使って下流のAUG部位に移動し、50S rRNAが結合して70S rRNAが形成される。
ペプチド鎖はtRNAとアミノ酸をつないでいる部分が切断され、アミノ酸同士のペプチド結合が形成されることでペプチド鎖が増えていく。
ペプチド鎖が増えていく部分をもう少し詳しく見てみようう。
50S rRNAの中にはP部位(ペプチジル部位)とA部位(アミノシル部位)が存在する。アミノ酸と結合したtRNAがコドンに結合するためにはeEF-1α(Elongation factor)とGTPが必要である。コドンに結合するということは、tRNAがA部位に入るということである。
A部位にtRNAが結合するとペプチジルトランスフェラーゼによってアミノ酸同士をペプチド結合でつなぎ、ペプチド鎖をA部位に移動させる。
P部位にあるtRNAがペプチド鎖をA部位に渡すと、tRNAはリボソームから解離する。その後、リボソームが動いてA部位だった部分がP部位となる。つまり、A部位が空となる。このようにリボソームが動き、tRNAがA部位からP部位へと移ることをトランスロケーションという。トランスロケーションにはeEF-2(Elongation factor 2)とGTPが必要である。
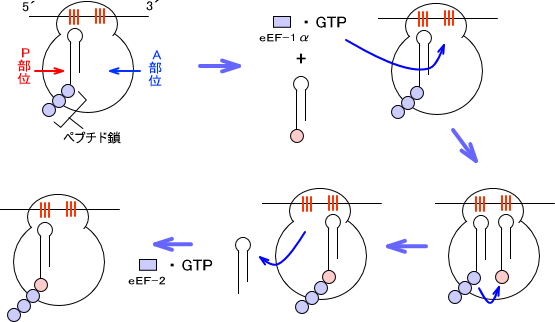
原核生物のリボソームは1秒間に18個のアミノ酸を取り込み、真核生物のリボソームは1秒間に6個のアミノ酸を取り込む。
また、翻訳を終結するとき終止コドンに対応するtRNAは存在しないため、遊離因子とGTPが行う。
翻訳過程
翻訳はN末端→C末端の方向で行われ、最初のメチオニンは切り離される。
翻訳が生じる場所は細菌は細胞質であり、真核細胞では細胞質or粗面小胞体上のリボソームで行われる。原核細胞でも真核細胞でも1本のmRNAには何個ものリボソームが結合しており、この状態をポリリボソームまたはポリソームという。
なお、リボソームから出てきたばかりのタンパク質はアミノ酸のヒモであるため、タンパク質として働くための適切な構造をとっていない。アミノ酸のヒモの状態のタンパク質は他の分子と結合しやすいなど、とても不安定である。
そのため、これらのタンパク質が適切な構造になるまで防御しないといけない。この働きをシャペロンが行う。
また、タンパク質が合成される最初の塩基(AUG)から停止コドンまでをオープンリーディングフレームという。
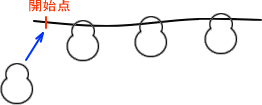
・ポリソーム
リボソームはmRNAの開始点に結合し、翻訳し始めると当然ながら翻訳のために移動していく。すると、他のリボソームが開始点に結合し、また翻訳して移動していく。
これが続くとmRNAには何個ものリボソームが結合している状態となる。この状態をポリソームという。
突然変異
・フレームシフト
mRNAに1個または複数の塩基が挿入・欠如することでmRNAの読み取りの枠(3個の組み合わせ)が変化することである。mRNAのコドンを正常に読むために句読点があればいいが、コドンを正常に読むための句読点はmRNAに存在しない。そのため、読み取り枠がずれて機能を失ったタンパク質が合成される。
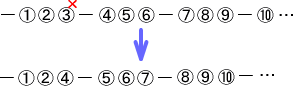
・ナンセンス変異(ペプチド鎖終結変異)
ナンセンス変異は「塩基が変化して終止コドンが生じる」という変異である。例えば「UCG」の「C」が「A」に変化すると終止コドンであるUAGとなる。
普通、UAGなどの終止コドンに対応するアンチコドンをもっているtRNAは存在しない。そのため、ナンセンス変異が生じるとペプチド鎖の合成が途中で中断し、機能をもたないタンパク質が合成される。
ただし、原核生物や下等な真核生物ではナンセンス変異が起きても、ペプチド鎖の合成が停止しないことがある。これはUAGなどの終止コドンに対応するアンチコドンをもつ異常なtRNAが存在するからである。このtRNAは3´末端にアミノ酸を有しており、ペプチド鎖をつなげる働きをする。
前述の通り、通常は終止コドンに対応するtRNAは存在しない。つまり、このようなtRNAはそれ自体が突然変異によって生まれたものである。このようにナンセンス変異を無効にするtRNAをサプレッサーtRNAという。
このようにサプレッサーtRNAはナンセンス変異を抑制する働きがあるが、正常な終止シグナルも抑制してしまうことがある。これにより、望ましくない読み過ごしが起きてしまう。
スポンサードリンク
カテゴリー
スポンサードリンク