遺伝子の変異
DNAの塩基の変異
・点変異
突然変異にはいくつかの種類がある。その中でも塩基対が一個変化した変異を点変異という。

AとGはプリン塩基でTとCはピリミジン塩基である。
点変異には「プリン塩基→プリン塩基 or ピリミジン塩基→ピリミジン塩基」の変化と「プリン塩基→ピリミジン塩基 or ピリミジン塩基→プリン塩基」の2種類の変化がある。前者をトランジションといい、後者をトランスバージョンという。
なぜこのように区別するかというと、プリン塩基とピリミジン塩基では構造が異なるからである。「プリン塩基からプリン塩基に変わる場合」と「プリン塩基からピリミジン塩基に変化する場合」では後者の方が変異による影響が大きい。
・欠損や挿入
DNAには塩基が挿入する場合やDNAから塩基が欠損する場合がある。
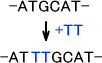
遺伝子レベルの変異
DNAの配列が変わるということはコードされたアミノ酸配列が変わるかもしれないということである。ただし、DNA配列が変わっても影響を与えない場合もあれば大きく影響を与えてしまう場合もある。
・静的変異(サイレント変異)
静的変異とはDNA配列には変化があるが、アミノ酸配列には無関係の場合の変異である。
例えば「CTA」はLeuをコードしていることを意味するが、C→Tに変化したとしてもLeuをコードする「TTA」に変化しただけなのでアミノ酸には全く影響を与えない。

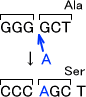
・ミスセンス変異
ミスセンス変異とはDNA配列が変化することによって、アミノ酸が置き換わることである。
例えば「TTA」はLeuのコードを意味するが、A→Tに変化すると「TTT」となりPheをコードすることになる。変異した場所のアミノ酸がタンパク質にとって重要でない部分ならさほど問題とならないが、変異した部分が重要な場所であればかなり問題である。

・ナンセンス変異
ナンセンス変異とはアミノ酸のコードが終止コドンに変化する変異のことである。
例えば「TTA」とコードしている配列があるとする。「TTA」はLeuのコードであるが、T→Gに変異すると「TGA」となり終止コドンへと変化する。
終止コドンに変化するとタンパク質の合成は途中でストップしてしまう。この場合、途中で途切れた短いタンパク質が合成されることになる。なお、このタンパク質のほとんどは活性がない。
・フレームシフト変異(読み枠移動変異)
塩基の挿入や欠損の結果として起こる変異である。この場合はアミノをコードする配列がすべて変化する。
自然に起こる遺伝子変異
別に発癌物質の作用がなくても遺伝子が自然に変異することはある。下に自然に起こる遺伝子変異を紹介する。
・ポリメラーゼの読み間違い
DNAの合成はDNAポリメラーゼが行う。しかし、このポリメラーゼが誤って塩基を挿入してしまったら変異が起きてしまう。
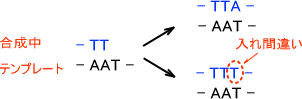
ポリメラーゼによる間違いは大腸菌で調べてみるとかなりの頻度で起きている。ポリメラーゼは10~100個の割合でミスがある。
ただし、ミスがあるとそれを修復するような機構が働く。この機構のために本当の意味でのエラーは10-4まで減少する。ポリメラーゼが挿入ミスをして、そのミスが修正されなかったら変異が起こってしまう。

・自然のフレームシフト変異
フレームシフトが自然に起こるときは同じ塩基がいくつも並んでいる場所で起こりやすい。塩基が1つ余分に挿入される場合は、合成中の鎖がずれて合成されるときに起こる。逆に塩基が1つ欠損する場合はテンプレートの方の鎖がずれて合成されるときに起こる。
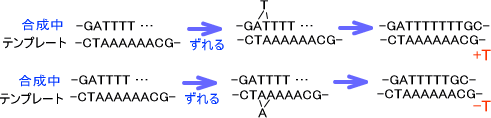
上の図の状態でもう一度複製が起こると、完全なフレームシフト変異が起きてしまう。
・塩基の互変異性
CはAと対を作る。しかしこれはCがケト型のときの場合であり、Cがエノール型のときはGと塩基対を形成してしまう。
これはポリメラーゼのミスでもないので仕方のない変異である。
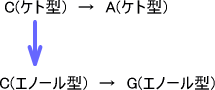
自然のDNAの変異
DNAは紫外線や発癌物質によって常に変異を受けている。つまり、決して「DNAは安定である」ということはできない。
自然に起こるDNAの変異の種類には下のようなものがある。
・脱塩基部位の生成
脱塩基は自然に起こる変異であり、常に発生している。
脱塩基ではデオキシリボースとプリンヌクレオチドを繋いでいるN-グリコシド結合が開裂する。これによって塩基が失われる反応を脱プリン反応という。プリン塩基にはアデニンとグアニンがある。
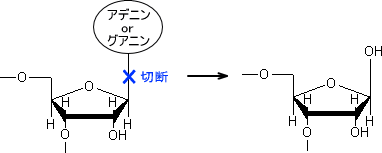
脱塩基が起こるということは、DNAには塩基が失われている場所が存在することになる。この部位をAPサイト(またはアベージックサイト)という。特にプリン塩基のAPサイトをアプリニックサイト、ピリミジン塩基のAPサイトをアピリミジニックサイトとい。
APサイトが修復されないうちにDNAポリメラーゼが来ると、DNAの複製は一端停止する。しかし、結局はAPサイトに適当な塩基を入れて先へ進む。これによって変異が起こる。なお、大腸菌はAPサイトにAを入れる性質がある。

・脱アミノ化
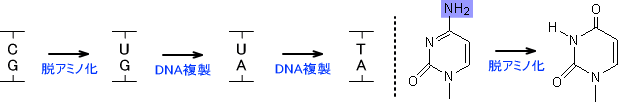
塩基からアミノ基が失われる反応である。この脱アミノ化は水によって起こる。シトシンが脱アミノ化するとウラシルに変化する。(C→U , 100塩基/日)
ウラシルはアデニンと対を作るので、脱アミノ化をそのままにしておくとDNAの複製によってU:G → U:Aとなり、もう一度複製を行うとU:A →
T:Aとなる。こうなるとC:Gが完全にT:Aとなる。
(C:G → U:G → U:A → T:A)
また、脱アミノ化の問題はシトシンだけでなく5-メチルシトシンにも存在する。通常のDNAのシトシンは約4%がメチル化されている。メチル化の結果、シトシンは5-メチルシトシンとなる。
5-メチルシトシンはシトシンと同じように脱アミノ化する。シトシンが脱アミノ化するとウラシルへと変化したが、5-メチルシトシンが脱アミノ化するとチミンへ変異する。この状態でDNAの複製が起こるとG:T → A:Tとなる。(G:MeC → G:T → A:T)
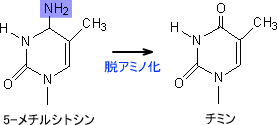
ウラシルはDNAには存在しないため変異であるとすぐに見分けることができるが、チミンはDNAに存在する正常塩基である。つまり、5-メチルシトシンの脱アミノ化は修復されにくい。修復される場合は親鎖と娘鎖の区別がつくときだけに限られる。このような理由で5-メチルシトシンは変異しやすい部分なのである。
また、DNA中での変異体数は一様に散らばっている。つまり、ほとんど同じ割合で変異が発生しているのである。しかし変異対数が通常の100倍にもなる部分があり、この場所をホットスポットという。
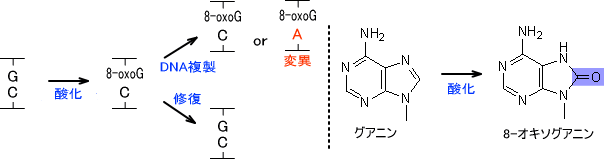
・塩基の酸化
DNAの塩基は酸化されると変異をもたらすことがある。チミンが酸化されるとチミングリコールとなり、グアニンが酸化されると8-オキソグアニン(8-ヒドロキシグアニン)に変化する。この8-オキソグアニンはアデニンともシトシンとも対合することができる。
誘導突然変異
自然に起こる突然変異もあれば、化学物質によって突然変異が誘導される場合がある。このような化学物質が発癌物質である。
・塩基類似物質
塩基に似ている物質がうろうろしているとDNAポリメラーゼが塩基と間違ってDNA上に組み込んでしまうことがある。これによって変異が起こる。
例えば、5-ブロモウラシル(BU)はアデニンともグアニンとも対合することができる。
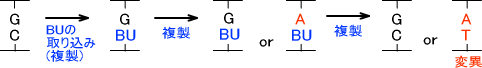
また、DNAの塩基は酸化が酸化されて8-オキソグアニンが生じ、これが変異の原因になることがあると前に紹介した。しかし、酸化されるのは塩基だけではない。塩基が挿入されるときは原料としてdNTP(デオキシヌクレオチド)が使われるが、この原料が既に酸化されている場合がある。
グアニンが挿入されるときはdGTPが原料となる。このdGTPが酸化されたd8oxoGTPの状態でDNAポリメラーゼによって挿入されるのである。
・メチル化
5-メチルシトシンは正常なメチル化であるが、正常でないメチル化も存在する。この正常でないメチル化にはO6-メチルグアニンなどがあり、O6-メチルグアニンは シトシンにもチミンにも対合する性質がある。

・脱塩基部位形成
これは化学物質によってAPサイトが形成される反応である。
例えば、ベンツピレン(B[a]P)の活性体がグアニンと結合すると脱塩基が起こる。これによってAPサイトが形成され、修復される前にポリメラーゼが来ると適当に塩基が入れられて変異が起こる。

・インターカレーションによるフレームシフト変異
この反応は化学物質によってフレームシフトが起こる変異である。
ベンツピレン(B[a]P)の活性体がグアニンと結合するとベンツピレンは塩基と塩基の間にはまり込む。塩基同士の間は狭いので、ベンツピレンは間を押し広げてしまう。
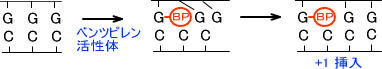
この状態で複製が行われると塩基が1つ余分に挿入され、フレームシフトが起きることがある。
スポンサードリンク
カテゴリー
スポンサードリンク