遺伝子発現の制御:転写制御とオペロン
遺伝子発現とはDNAの情報がmRNAに転写され、mRNAの情報を翻訳してタンパク質を合成する過程である。転写と翻訳には多くのエネルギーを必要とする。そのため、タンパク質は必要なときだけに合成される必要があり、遺伝子の発現は制御されないといけない。
導入部分
それでは簡単な質問を二つしようと思う。○か×で答えよ。
問1 遺伝子は体の部分によって特別な遺伝子(脳なら脳のためだけの遺伝子など)をもつ。
問2 映画「ジュラシック・パーク」のようなDNA(遺伝子)だけで、製品(恐竜)を作れる。
できれば質問に答えるだけでなく、なぜそのような答えになるのかも考えてほしい。それでは下に質問の答えを記す。
答1 ×
それでは、なぜ「脳は脳だけの遺伝子、心臓は心臓だけの遺伝子をもたない」と言えるのだろうか。クローン羊ドリーの話を思い出してほしい。ドリーは体細胞を使ったクローンである。
体細胞は髪の毛の細胞でも皮膚細胞でもどこでもいい。どの部分でも同じ遺伝子をもつので、クローンではどの体細胞を使ってもかまわないのである。
ただし、「遺伝子は体のどの部分でも同じ遺伝子をもつ」というのは誤りである。ヒトの全ての遺伝子で抗体を産出するリンパ球だけは他の体細胞と異なる遺伝子セットをもつのである。分化した体細胞からクローンが作れるが、リンパ球だけは例外である。
答2 ×
現在の科学では不可能である。
DNAからRNAが作られ、RNAからタンパク質が作られる。タンパク質が機能するには三次構造をとる必要がある。つまり、タンパク質が適切な構造をとらなければ機能しなかったり、機能が異なったりするのである。
DNAは決められた状態で決められた手順に基づいて発現し機能する必要がある。DNAだけでは発現制御できないのである。
それでは、クローン羊ドリーはどのようにして作られたのだろうか。それは、体細胞からのDNAを他の羊の卵細胞に挿入する方法である。同じ羊の卵細胞なら適切に遺伝子発現してもおかしくないはずである。
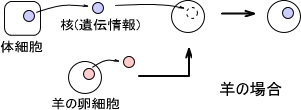
問題はトカゲなどの生物の卵細胞に恐竜の遺伝子を挿入して恐竜が生まれるかである。こう考えると、まず不可能であろう。
転写制御
遺伝子の制御は負の制御と正の制御の二種に分けることができる。
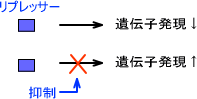
負の制御をするタンパク質がDNAに結合すると、特定の遺伝子の転写を妨げる。このような働きをするタンパク質をリプレッサー(負の制御因子)という。
逆に正の制御するタンパク質がDNAに結合すると、特定の遺伝子の転写を活性化する。このような働きをするタンパク質をアクティベーター(正の制御因子)という。
負の調節 |
正の調節 |
|
調節因子存在 |
減少 |
増加 |
調節因子なし |
増加 |
減少 |
遺伝子にリプレッサーが結合している状態では転写が妨げられている。転写が開始させるためにはリプレッサーを取り除かなくてはならない。リプレッサーを取り除く働きをする分子を誘導物質(inducer)という。

リプレッサーの発現を抑えると結果として遺伝子発現が増える。つまり、このようなエフェクターはアクティベーターのように見ることができる。
シグナル応答
リプレッサーやアクティベーターはシグナル分子と結合することで応答するものもある。シグナル分子は時間的応答に従い、三種に分けることができる。
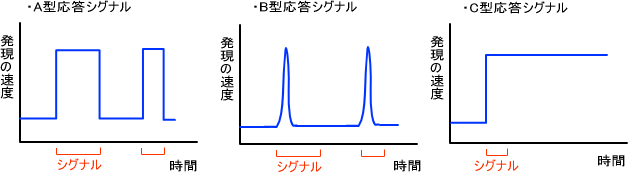
A型応答形式:誘導シグナル(ホルモンなど)に応じて発現
発現速度は誘導シグナルが存在することに依存している。誘導シグナルが存在すれば調節因子が機能を示す。つまり、発現速度が増加または減少する。これらの誘導シグナルが存在しなければ基底状態に戻る。
B型応答形式:一過性に発現
誘導シグナルが継続して存在していても一過性にしか発現しない。シグナルがなくなり、細胞が回復してからシグナルが与えられるとまた一過性に発現する。
C型応答形式:調節シグナル後も発現
誘導シグナルが消失しても遺伝子発現の速度が元の状態に戻らないのがこのパターンである。このとき、シグナルは引き金(trigger)としての役割をしている。この応答形式の典型的な例は分化のときの過程である。
遺伝子発現
遺伝子(DNA) → mRNA → タンパク質
このような場合、「遺伝子が発現している」ということができる。
大腸菌の場合、mRNAの合成量はタンパク質合成量にほぼ比例している。これは、大腸菌が転写レベルで調節しているためである。しかし、真核生物では複雑な制御系が存在するため、転写レベルでは調節されていない。
原核生物、真核生物ともに細胞が生存し増殖するために必要なハウスキーピング遺伝子がある。ハウスキーピング遺伝子は多くの細胞に存在し、一定量だけ発現する。これらの遺伝子は例えばタンパク質産出などの生きていくために必要な情報を有している。
原核生物の遺伝子
mRNAが転写されるとき、DNAの一つのプロモーターから一本の連続したmRNAができる。このmRNAから各々のタンパク質に翻訳される。このような転写の制御単位をオペロンという。
一種類のタンパク質をコードする遺伝形質の最小単位をシストロンという。また、一本のmRNAから複数のタンパク質が合成されるmRNAをポリシストロニックmRNAという。一本のmRNAにシストロンが複数存在するのである。なお、ポリシストロニックmRNAは真核細胞では見られない。
ラクトースオペロン(lacオペロン)
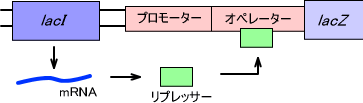
ラクトースオペロンは大腸菌のラクトース分解に関わる酵素の遺伝子群である。ラクトースオペロンにはラクトースオペロン制御に関するlacI (リプレッサー)、プロモーター、オペレーターとラクトース代謝酵素をコードするlacZ (β-ガラクトシダーゼ)、lacY (ラクトース輸送体)、lacA (チオガラクトシドトランスアセチラーゼ)が存在する。
lacZ はラクトースの分解酵素、lacY はラクトースを細胞内へ通過させるための透過酵素、lacA はラクトースを分解したときに産出するガラクトースの分解酵素をコードしている。
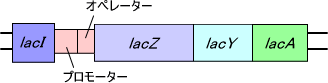
lacI は常に発現しており、一定速度でlacリプレッサーを合成している。このようにlacI は構成的に発現している。オペレーター(O領域)は遺伝子発現制御に関しており、lacI で合成されたリプレッサーがオペレーター領域に結合することでlacオペロンの発現が抑制される。
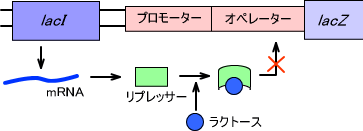
ラクトースが存在するとlacY を介して細胞内に取り込まれ、リプレッサーに結合する。ラクトースがリプレッサーに結合すると、リプレッサーの形が変わって不活性型になりlacI による抑制が解除される。これによりlacZ -lacY -lacA の発現が開始される。
カタボライトリプレッション(異化代謝産物抑制)
大腸菌などでは培地中にグルコースが十分にあるとき、ラクトース、ガラクトース、アラビノースなどの他の糖が高濃度に存在していても、その発酵に関わるタンパク質は作られない。
これは、グルコースの方が使いやすい糖であるためであり、エネルギーの無駄遣いを防ぐためである。
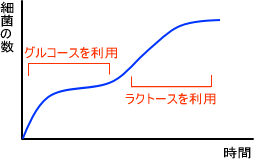
上の図は二段増殖といい、大腸菌をグルコースとラクトースで培養したときの時間と増殖数の関係である。最初はグルコースを利用し、グルコースが無くなるまではラクトース利用系の発現が抑制されている。
また、lacオペロンのプロモーター領域にはRNAポリメラーゼが結合する領域の他にCAP結合部位という領域が存在する。CAPはcAMPと複合体をつくり、CAP結合部位に結合する。
CAP-cAMP複合体はlac遺伝子の制御に関わっており、cAMP-CAP複合体がCAP結合部位に結合することで、ポリメラーゼがプロモーターに結合しやすくなる。 →lacオペロンの発現が促進される。
cAMPはアデニル酸シクラーゼという酵素によって合成される。しかし、グルコースはアデニル酸シクラーゼを阻害するためcAMPが合成されない。cAMPの濃度が下がるためCAP-cAMP複合体ができず、プロモーターに結合することができなくなり発現は抑制される。
CAPはCRPやcAMP調節タンパク質とも呼ばれている。
lacオペロンの発現には次の条件が必要である
・誘導物質であるラクトースや無償(使われない)の誘導物質が存在する。
・cAMP濃度が高い(グルコースレベルの低下)
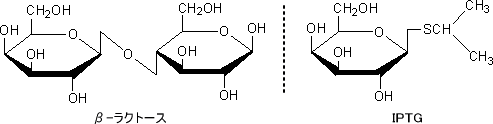
無償の誘導物質とは「リプレッサーに結合して発現を促進(誘導)するが、β-ガラクトシダーゼによって分解されない(グルコースを産出しない)物質」のことである。例としてIPTGなどのガラクトースなどの構造類似体がある。
cAMP(サイクリックAMP)は多くの生物に存在し、アデニル酸シクラーゼによって合成される。グルコースはアデニル酸シクラーゼの合成を阻害する働きがあるため、cAMPの濃度はグルコース濃度に左右される。つまりグルコース濃度が高いとcAMP濃度は低く、グルコース濃度が低いとcAMP濃度は高い。
アラビノースオペロン(正の調節)
基本的にラクトースオペロンと似ている。アラビノース存在下で発現促進し、カタボライトリプレッションを受ける。
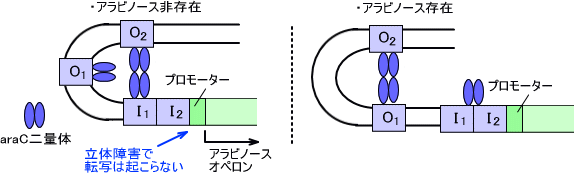
アラビノースオペロンの発現にはaraCというタンパク質が必要になる。ただし、araCは条件によってはリプレッサーとしてもアクティベーターとしても働く。アラビノースが存在するならaraCはアクティベーターとして働き、アラビノースが存在しないならリプレッサーとして働く。
araCがリプレッサーとして働くときはaraC二量体がO1とI1-O2に結合し、立体障害によってアラビノースオペロンの発現が抑えられている。
アラビノース存在下ではaraCタンパク質二量体にアラビノースが結合する。この複合体はI1-O2ループをほどき、I1,I2と結合して転写を活性化させる。
トリプトファンオペロン
大腸菌にとってトリプトファンは不可欠であり、必要なときは自分で生合成する。ただし細胞内にトリプトファンが十分にあれば発現は止まり、ラクトースオペロンと逆の作用を示す。つまり、トリプトファンがあるときは生合成が抑制される。
過剰のトリプトファンはトリプトファンリプレッサーと結合する。その結果、トリプトファンリプレッサーは不活性型から活性型に変わり、オペレーターに結合してトリプトファンの生合成を抑制する。(Trpがなくなると合成が再開する)
レギュロン
遺伝子発現の制御では「正の制御」「負の制御」に関わらず、制御タンパクは高い特異性をもつことが多い。例えば、lacI (リプレッサー)はラクトースオペロンのみに働く。
ただし、一種類の制御タンパク質が広範囲に制御することがある。このとき、一種類のタンパクによって広範囲に制御される遺伝子群やオペロン群をレギュロンという。
スポンサードリンク
カテゴリー
スポンサードリンク